Blood Cell Differentiation, Maturation
and Function
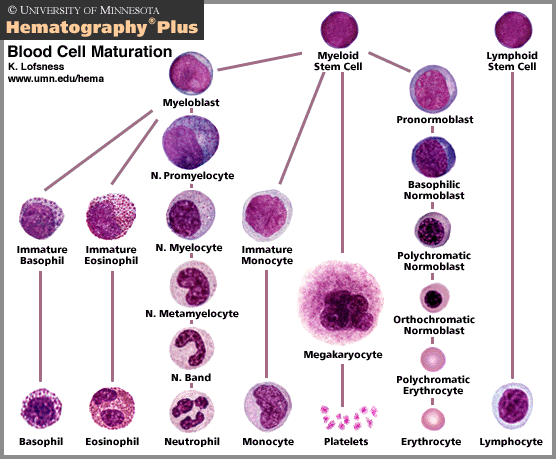
Red cells, white cells and platelets are made in the marrow of bones, especially the vertebrae, ribs, hips, skull and
sternum. These essential blood cells fight infection, carry oxygen and help control bleeding.
Mature cells are usually specialized. As cells mature, their shape and contents change so that they can be most efficient at the work that they must do. All of the mature blood cells come from one type of precursor, a stem cell.
Not all cells are capable of dividing to make new cells. Cells start from a "parent" or stem cell, but then, under instructions from the rest of the body, they begin to specialize and mature. They may alter almost anything and everything about themselves to get ready to do a specific job or set of jobs. They may change shape, their nucleus may be discarded, they may attach to other cells or shed little pieces of themselves. They may make and secrete certain substances or make the substances and store them until they are to be used.
In the blood, all of the white blood cells keep their nuclei so that they can alter their function to meet the challenges of protecting the body from invaders. The red cells, however, lose their nuclei and fill up with the special oxygen-carrying molecule, hemoglobin. They also assume a special shape that is flexible, yet allows rapid transfer of oxygen into and out of the cell.
The megakaryocyte stays in the bone marrow after it differentiates from the stem cell. Megakaryocytes are like factories. They manufacture and shed millions of little pieces of themselves into the blood. These pieces of cell, called platelets, contain substances necessary to help blood clot.
CELLS AND ORGANS OF THE LYMPHOID SYSTEM
All leukocytes originate in the bone marrow from self-renewing hemopoietic stem cells.
These stem cells give rise to four major cell lineages: erythroid (erythrocytes),
megakaryocytic (platelets), myeloid (granulocytes and monocytes/macrophages), and lymphoid
(T and B lymphocytes and natural killer cells).
CELL LINEAGES. All leukocytes are derived from pluripotent stem cells in the bone marrow
and liver which give rise to two main progenitors; one for lymphoid cells and the other for
myeloid cells. The common lymphoid progenitor has the capacity to differentiate into either
T or B cells depending on the microenvironment to which it homes. In mammals, T cells
develop in the thymus while B cells develop in the bone marrow. The precise origin of some
antigen-presenting cells (e.g. dendritic cells) is uncertain, although they do develop
ultimately from the hemopoietic stem cells. NK cells also derive from the common lymphoid
progenitor cell. The myeloid cells differentiate into the committed cells shown on the left
in the diagram above.
Lymphoid Lineage
109 lymphocytes are produced each day by the human body. The average adult has about 1012
total lymphocytes, and lymphoid tissue as a whole accounts for about 2% of body weight.
Lymphocytes comprise 20% of all leukocytes.
Lymphocytes range in size from 6-10 mm diameter and are relatively indistinguishable by
light microscopy. However, the cells express a large number of different protein "markers"
on their surfaces that can be used to distinguish among the populations. The presence of
the markers can be detected using specific monoclonal antibodies that bind to them. A
systematic nomenclature (the "cluster of differentiation/designation" or "CD" system) has
been developed to identify the markers. One marker that all leukocytes have in common (a
pan-leukocyte marker) is CD45.
I. T cells
T lymphocytes arise in the bone marrow, then migrate to and mature in the thymus. All T
cells possess the T cell antigen receptor (TCR), which serves as the T cell's definitive
marker. In addition, all T cells possess the CD3 marker, a complex of 5 polypeptides
thought to be involved in transmembrane signal transduction. There are two major types of T
cells, helper and cytotoxic T cells.
A. Helper T cells (TH) secrete cytokines that promote the proliferation
and differentiation of T cells, B cells and macrophages, and that recruit
and activate inflammatory leukocytes. TH cells are identified by the
presence of the CD4 marker. TH cells are further subdivided into the
TH1 and TH2 subsets on the basis of the kinds of cytokines they
produce. Both subsets are derived from a precursor known as TH0,
which produces all cytokines made by either subset (listed below) and
interleukin-3 (IL-3).
1. TH1 (inflammatory helper cells)
TH1 cells produce interleukin-2 (IL-2), interferon-gamma (IFNg), and
tumor necrosis factor-beta (TNFb), cytokines that promote cellular
immune responses.
2. TH2 (helper cells)
TH2 cells produce IL-4, IL-5, IL-6, IL-10 and TGFb, all of which
promote antibody synthesis by B cells.
B. Cytotoxic or cytolytic T cells (Tc) lyse cells that produce foreign
antigens, e.g. tumor cells, virus-infected cells, and foreign tissue grafts.
Tc cells are identified by the presence of the CD8 marker.
II. B Cells
B lymphocytes acquired their name because in birds they were first shown
to mature in an organ called the Bursa of Fabricius. In mammals, the early
stages of B cell maturation occur in the fetal liver and bone marrow.
Surface immunoglobulin (sIg) is the B cell's receptor for antigen and its
definitive marker. Two accessory molecules, Iga and Igb, are involved in
B cell activation, just as CD3 participates in T cell activation. Additional
markers that distinguish B cells include CD19, CD20, CD21 and CD22. B
cells may differentiate into plasma cells, which secrete large amounts of
antibodies, or into memory B cells. Memory cells can survive 20 years or
more in the absence of antigen. Upon secondary contact with antigen,
they proliferate rapidly and begin active antibody secretion. (T cells can
also form memory cells).
III. Natural killer (NK) cells
NK cells are lymphocytes that do not express either sIg or TCR. All NK
cells express CD16, a receptor for a portion of the IgG antibody molecule
called the Fc fragment. CD16 allows the NK cell to kill target cells coated
with IgG. NK cells are also important in eliminating tumor cells and
virus-infected cells.
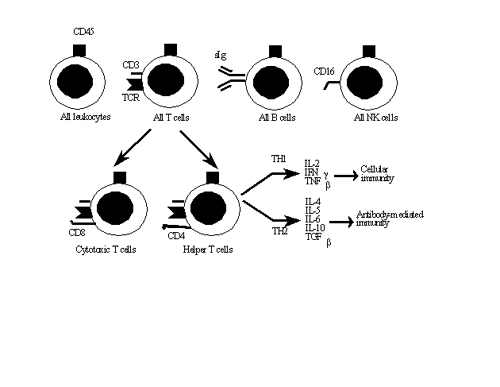
The important lymphocyte markers are indicated in the illustration above.
Myeloid Lineage
Broadly, there are two major types of myeloid cells, the mononuclear phagocytes (monocytes
and macrophages) and the granulocytes (neutrophils, eosinophils, and mast cells/basophils).
Differentiation of the myeloid progenitor into each committed cell type depends upon which
interleukins and colony stimulating factors are present in the milieu where the cells are
maturing. Sources of the ILs and CSFs include stromal cells and mature myeloid and lymphoid
cells.
I. Mononuclear phagocytes (10-18 �m diameter)
Monocytes and macrophages can remain viable for months or years. The
cells fall into two categories, the professional phagocyte, whose major
function is to remove particulate antigens, and the antigen-presenting cell,
whose function is to present antigen to lymphocytes.
A. Professional phagocytes
Monocytes/macrophages adhere to tumor cells or microorganisms
through mannosyl-fucosyl receptors that bind to carbohydrates, or
through receptors for complement or IgG. This binding may trigger
extracellular killing, phagocytosis, or both, of the target cells.
Monocytes/macrophages produce hydrolytic enzymes, reactive oxygen
metabolites (e.g. H2O2, superoxide anions, singlet oxygen and hydroxyl
radicals), complement components, tumor necrosis factor a, and IL-1,
all of which are toxic to tumor cells and microorganisms.
B. Antigen-presenting cells (APCs)
The TCR on TH cells cannot interact with antigen unless the antigen is
bound to the surface of an antigen-presenting cell. Besides
macrophages, many other kinds of cells can act as APCs, including B
lymphocytes, dendritic cells and the Langerhans cells of the skin. APCs
internalize large antigens, digest the antigens into small fragments, and
then export the fragments to the cell surface. Interaction of the TH cell
with the small fragments on the surface of the APC causes the TH cells
to produce cytokines that stimulate antibody synthesis in B cells or
cytotoxicity in TC and NK cells.
II. Polymorphonuclear granulocytes
Polymorphs are the most prevalent of the leukocytes, being released from the
bone marrow at the rate of 7 million/min. These cells live 2-3 days.
A. Neutrophils are characterized by their multilobed nuclei. Their primary
function is phagocytosis, which is enhanced by opsonization with
complement and antibodies. An important secondary function is to
promote inflammation. Like the mononuclear cells, neutrophils produce
reactive oxygen metabolites and hydrolytic enzymes. In addition,
neutrophils produce nitric oxide and antibiotic proteins such as
defensins, seprocidins, cathelicidins, and bacterial permeability inducing
protein.
Lysosomes in neutrophils contain granules of two types. Primary
(azurophilic) granules contain acid hydrolases, myeloperoxidase and
lysozyme. The secondary, or specific, granules contain lysozyme and
lactoferrin.
B. Eosinophils express low affinity Fc receptors specific for IgE and are
important in reactions to antigens (e.g. parasites) that induce high levels
of IgE. Eosinophils are particularly effective at destroying helminthic
parasites, which are often resistant to the lysosomal enzymes of
neutrophils and macrophages. The granules in eosinophils release major
basic protein and eosinophil cationic protein, which are toxic to the
worm, and substances like histaminase and arylsulfatase that reduce
inflammation.
C. Basophils and mast cells are the least prevalent of the leukocytes. They
possess high affinity Fc receptors for IgE and release the chemical
mediators of immediate hypersensitivity, including histamine,
prostaglandins, thromboxanes, leukotrienes, and heparin. They also
produce eosinophil chemotactic factor (ECF) which causes eosinophils
to enter the area of worm infestation or allergen localization.
Differential White Blood Cell Count
CELL TYPE % OF WBC'S CELLS/cmm
Neutrophil 50 - 60 3000 - 7000
Eosinophil 1 - 4 50 - 400
Basophil 0.5 - 2 25 - 100
Lymphocyte 20 - 40 1000 - 4000
T cell 80 - 85*
B cell 5 - 15*
NK cell 5 - 15*
Monocyte 2 - 9 100 - 600
*% of lymphocytes in peripheral blood
Megakaryocytic Lineage
Platelets, which are anucleate, are derived from megakaryocytes and contain granules at the
ultrastructural level. Their major functions are blood clotting and inflammation.
Following injury to endothelial cells, platelets adhere to the surface of the damaged
tissue, where they release substances that increase vascular permeability, activate
complement and attract leukocytes.
Lymphoid Organs
The lymphoid organs are those organs in which maturation, differentiation and proliferation
of lymphocytes take place. The primary, or central, lymphoid organs are those in which T
and B lymphocytes begin expressing their antigen receptors. The secondary lymphoid organs
are those in which lymphocytes are induced to proliferate and differentiate by contact with
antigen.
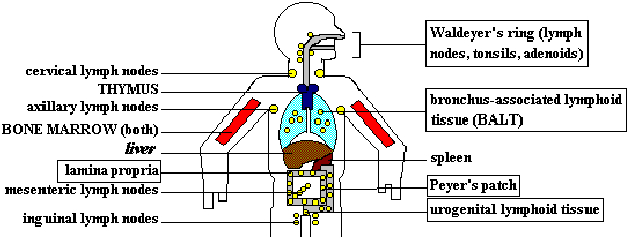
LYMPHOID ORGANS. The THYMUS and BONE MARROW are primary lymphoid organs. They are the sites
of maturation for T and B cells, respectively. Cellular and humoral immune responses occur
in the secondary (peripheral) lymphoid organs and tissues; effector and memory cells are
generated here. Secondary lymphoid organs can be classified according to the body regions
which they defend. The spleen responds predominantly to blood-borne antigens. Lymph nodes
mount immune responses to antigens circulating in the lymph, absorbed either through the
skin (superficial nodes) or from internal viscera (deep lymph nodes). Tonsils, Peyer's
patches and other mucosa-associated lymphoid tissue (MALT, boxes) respond to antigens which
have penetrated the mucosal barriers. Note that bone marrow is both a primary and secondary
lymphoid organ. Recently, it has become accepted that the liver is also a hematopoietic
organ, giving rise to all leukocyte lineages.
I. Primary (central) lymphoid organs
There are two major primary lymphoid organs, one in which T cells develop
(thymus), and the other in which B cells develop (bone marrow). The
structure of the thymus is presented below.
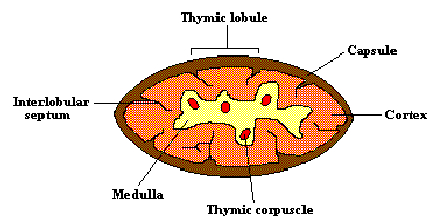
Progenitor cells from the bone marrow migrate to the thymus gland to
develop into T lymphocytes. The thymus is a bilobed structure, whose size
reaches its maximum at birth, then atrophies with age. The cortex contains
mostly immature thymocytes, some of which mature and migrate to the
medulla, where they learn to discriminate between self and nonself during
fetal development and for a short time after birth. T cells leave the medulla
to enter the peripheral blood circulation, through which they are transported
to the secondary lymphoid organs. It is in these secondary lymphoid organs
that the T cells encounter and respond to foreign antigens.
Removal of the thymus from an adult generally has little immediate effect on
the number and quality of the T lymphocytes, which have already matured
and populated the secondary lymphoid organs. In contrast, congenital
absence of a thymus results in an immediate and drastic reduction in T cells
that produces a potentially lethal wasting disease (DiGeorge syndrome).
Patients who survive the immediate neonatal period are prone to developing
recurrent or chronic infections with viral, bacterial, fungal or protozoal agents.
II. Secondary lymphoid organs
The secondary lymphoid organs have two major functions: they trap and
concentrate foreign substances, and they are the main sites of production of
antibodies and antigen-specific T cells. The major secondary lymphoid
organs include the spleen, which is responsive to blood-borne antigens; the
lymph nodes, which protect the body from antigens that come from skin or
internal surfaces via the lymphatic system; and the mucosa-associated
lymphoid tissue (MALT) scattered along mucosal linings, which protects the
body from antigens entering the body directly through mucosal surfaces.
A. Spleen
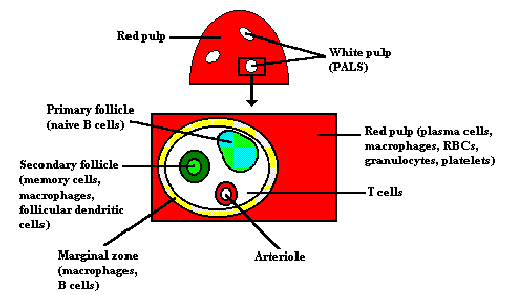
Schematic representation of the spleen and a cross-section of the periarteriolar lymphoid
sheath (PALS).
The spleen is the major organ in which antibodies are synthesized and
released into circulation. The spleen is composed of two types of tissue-the
red pulp and the white pulp. The red pulp contains plasma cells, resident
macrophages, erythrocytes, platelets, granulocytes and lymphocytes. It is the
site where aged platelets and erythrocytes are destroyed (hemocatheresis).
The white pulp contains the lymphoid tissue clustered around a central
arteriole in an arrangement known as a periarteriolar lymphoid sheath
(PALS). T cells are found around the central arteriole, while B cells are
organized into either primary follicles (unstimulated), or secondary follicles
(stimulated by contact with antigen) that contain a germinal center complete
with memory cells. About 50% of spleen cells are B lymphocytes and
30-40% are T cells.
Arterial blood lymphocytes enter the spleen through the trabecular artery and
return to venous circulation through the trabecular vein.
B. Lymph Node
Clusters of lymph nodes (ovoid structures usually less than 1 cm in
diameter) are strategically placed in the neck, axillae, groin,
mediastinum and abdominal cavity, where they act to filter antigens
from the interstitial tissue fluid and the lymph during its passage from
the periphery to the thoracic duct. Lymph nodes that protect the skin
are termed somatic nodes, while deep lymph nodes protecting the
respiratory, digestive and genitourinary tracts are termed visceral nodes.
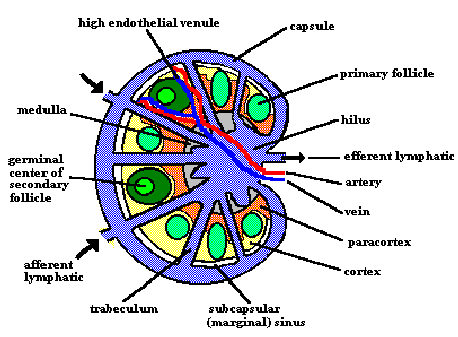
Schematic structure of the lymph node.
The lymph node consists of a B cell area (cortex), a T cell area
(paracortex) and a central medulla, which contains a mix of B cells, T
cells, plasma cells and macrophages. As in the spleen, B cells are
arranged in unstimulated primary follicles, and in antigen-stimulated
secondary follicles with germinal centers.
Antigen and lymphocytes enter the lymph node through the afferent
lymphatic vessels and high endothelial venules (HEVs). Lymph,
antibodies and cells leave the node through the efferent lymphatic
vessel. As the lymph passes across the nodes from afferent to efferent
lymphatic vessels, particulate antigens are removed by phagocytic cells
and are transported to the interior of the node where they stimulate
responses by B and T lymphocytes. Lymphoid cells return to
circulation via the thoracic duct, which empties into the left subclavian
vein.
C. Mucosa-associated lymphoid tissue (MALT)
The bulk of the body's lymphoid tissue (>50%) is found associated with
the mucosal system. MALT is composed of gut-associated lymphoid
tissues (GALT) lining the intestinal tract, bronchus-associated lymphoid
tissue (BALT) lining the respiratory tract, and lymphoid tissue lining the
genitourinary tract. The major effector mechanism at these sites is
secretory immunoglobulin A (sIgA) secreted directly onto the mucosal
epithelial surfaces. Examples of MALT include the Peyer's patches
lining the small intestine, the tonsils and the appendix.
Diffuse accumulations of lymphoid tissue are seen in the lamina propria
of the intestinal wall. The intestinal epithelium overlying the Peyer's
patches is specialized to allow the transport of antigens into the
lymphoid tissue. This particular function is carried out by epithelial
cells termed "M" cells, so called because they have numerous microfolds on
their luminal surface. M cells absorb, transport, process and present
antigens to subepithelial lymphoid cells.
III. Influence of adhesion molecules on lymphocyte traffic
1-2% of all lymphocytes recirculate each hour, allowing a large number of
antigen-specific lymphocytes to come into contact with the appropriate
antigen within the peripheral lymphoid organs. Lymphocyte traffic is
directed by the cuboidal endothelial cells lining the high endothelial venules
of the lymph nodes. When activated, the endothelial cells express adhesion
molecules that act as a kind of "glue" for the lymphocytes, preferentially
retaining and recruiting lymphocytes to the site of antigen localization. The
adhesion molecules expressed by MALT differ from those expressed by
systemic lymphoid organs. Thus, lymphoid cells stimulated in the MALT
tend to "home" back to the MALT, while systemic lymphocytes stay in the
periphery. Furthermore, adhesion molecules expressed by the HEVs differ
from those expressed at sites of inflammation. The adhesion molecules fall
into four families of structurally-related molecules-the immunoglobulin
supergene family, integrins, selectins and carbohydrate ligands.
SUMMARY
1. All leukocytes originate in the bone marrow and liver.
2. Lymphoid cells include B and T lymphocytes, plus NK cells.
3. B cells mature in the bone marrow. The definitive marker for the B cell
is surface immunoglobulin (antibody).
4. T cells mature in the thymus. The definitive marker for the T cell is the T cell
receptor for antigen (TCR).
5. TH1 cells promote cellular immune responses. TH2 cells help B cells make
antibodies. Both kinds of TH cells possess the CD4 marker.
6. TC cells kill tumor and virus-infected cells and foreign tissue grafts. They
bear the CD8 marker.
7. NK cells kill IgG-coated target cells, tumors and virally infected cells.
8. Myeloid cells include the mononuclear phagocytes (macrophages, monocytes)
and granulocytes (neutrophils, eosinophils, basophils, mast cells).
9. The major functions of the macrophage are phagocytosis and antigen
presentation. Macrophages kill engulfed microbes by producing hydrolytic
enzymes, oxygen radicals and cytokines.
10. The major function of the neutrophil is phagocytosis. Neutrophils kill
engulfed microbes by producing reactive oxygen metabolites, hydrolytic
enzymes, nitric oxide and cationic proteins.
11. The major function of the eosinophil is destruction of parasites in conjunction
with IgE.
12. Basophils and mast cells cause immediate hypersensitivity reactions.
13. Platelets (of megakaryocytic lineage) promote blood clotting and
inflammation.
14. The primary lymphoid organs (bone marrow and thymus) are the sites where
lymphocytes acquire their antigen receptors.
15. The thymus is composed of a cortex (immature cells) and a medulla (mature
cells). The organ atrophies with age.
16. The secondary lymphoid organs include the spleen, lymph nodes, and MALT
(tonsils, appendix, Peyer's patches). Secondary lymphoid organs serve to
trap antigens, allow antigen-specific cells to proliferate, and function as the
main sites of antibody production.
17. Most antibodies are made in the spleen. The organ is composed of red pulp
and white pulp. The white pulp contains most of the B and T lymphocytes,
arranged in periarteriolar lymphoid sheaths.
18. Primary follicles of the spleen contain lymphocytes that have not yet been
stimulated by antigen. Secondary follicles have been antigen-stimulated; they
contain germinal centers which function in B cell memory.
19. Lymph nodes are composed of a cortex (B cells), paracortex (T cells) and
medulla (mixture). The cortex contains both primary and secondary
lymphoid follicles.
20. Adhesion molecules expressed by endothelial cells lining the high endothelial
venules (HEVs) of the lymph nodes serve to attract and retain lymphocytes at
sites of antigen localization. They fall into 4 families: immunoglobulin
supergene family, integrins, selectins and carbohydrate ligands.
21. Lymphocytes within the MALT tend to home back to the MALT, while
systemic lymphocytes recirculate among the peripheral lymph nodes.
Reference Sources
1) Immunology, Kirksville College of Osteopathic Medicine; Kirksville, Missouri
2) www.umn.edu/hema
|

 E-mail
Irving
E-mail
Irving